
Virological and Parasitological Characterization of Mini-LEWE Minipigs Using Improved Screening Methods and an Overview of Data on Various Minipig Breeds

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mini-LEWE: Genetics and Breeding Conditions
2.2. Samples and Mitogen Stimulation of Peripheral Blood Mononuclear Cells
2.3. DNA and RNA Isolation
2.4. PCR and Real-Time RT-PCR for the Detection of Porcine Endogenous Retroviruses
2.5. Real-Time RT-PCR for the Detection of Hepatitis E Virus
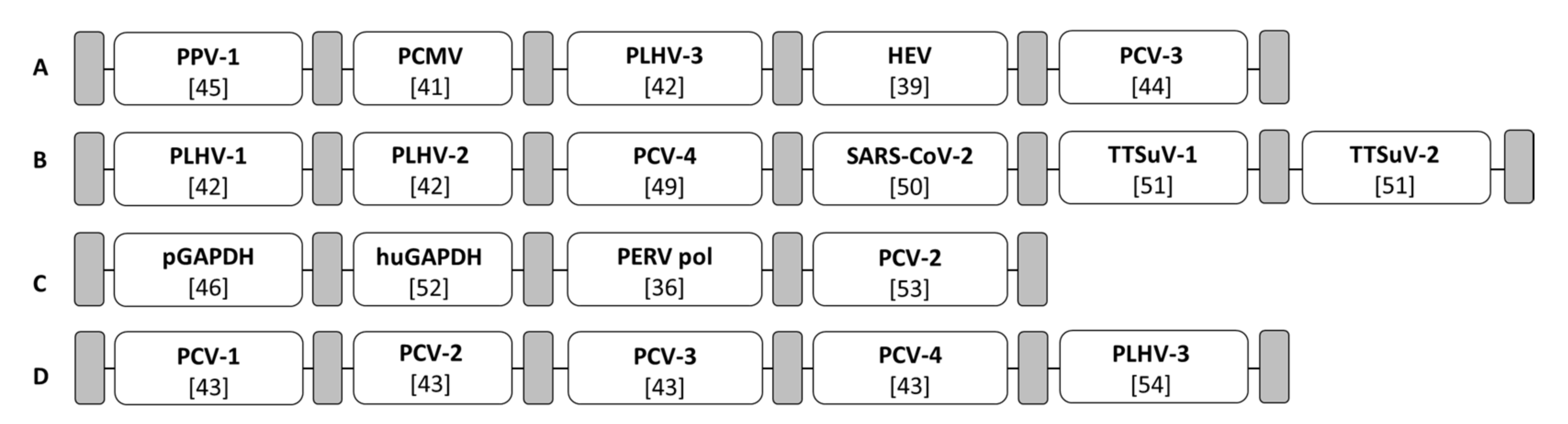
2.6. PCR and Real-Time PCR for the Detection of DNA Viruses
2.7. Generation of Viral Standards
2.8. Minimum Information for Publication of Quantitative Real-Time PCR Experiments (MIQE)
2.9. Parasitological Screening
3. Results
3.1. Prevalence of Porcine Endogenous Retroviruses
3.2. Screening for Hepatitis E Virus
3.3. Screening for Porcine Herpes Viruses, Circoviruses and Porcine Parvovirus-1
3.4. Establishment of gBlocks for the Standardization
3.5. Screening for Pig Parasites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 13, 5758. [Google Scholar] [CrossRef] [PubMed]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef] [PubMed]
- McAnulty, P.A.; Dayan, A.D.; Ganderup, N.-C.; Hastings, K.L. (Eds.) The Minipig in Biomedical Research; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Bollen, P.J.A.; Hansen, A.K.; Alstrup, A.K.O. The Laboratory Swine, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Gutierrez, K.; Dicks, N.; Glanzner, W.G.; Agellon, L.B.; Bordignon, V. Efficacy of the porcine species in biomedical research. Front. Genet. 2015, 6, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panepinto, L.M. Miniature Swine Breeds used Worldwide in Research. In Advances in Swine in Biomedical Research; Springer: Singapore, 1996; pp. 681–691. [Google Scholar]
- Panepinto, L.M.; Kroc, R.L. History, genetic origins, and care of Yucatan miniature and micro pigs. Lab. Anim. 1995, 24, 31–34. [Google Scholar]
- Ellegaard Göttingen Minipigs. Available online: https://minipigs.dk/goettingen-minipigs (accessed on 8 July 2021).
- Van der Windt, D.J.; Bottino, R.; Kumar, G.; Wijkstrom, M.; Hara, H.; Ezzelarab, M.; Ekser, B.; Phelps, C.; Murase, N.; Casu, A.; et al. Clinical islet xenotransplantation: How close are we? Diabetes 2012, 61, 3046–3055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morozov, V.A.; Ludwig, S.; Ludwig, B.; Rotem, A.; Barkai, U.; Bornstein, S.R.; Denner, J. Islet cell transplantation from Göttingen minipigs to cynomolgus monkeys: Analysis of virus safety. Xenotransplantation 2016, 23, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Ekser, B.; Ezzelarab, M.; Hara, H.; van der Windt, D.J.; Wijkstrom, M.; Bottino, R.; Trucco, M.; Cooper, D.K. Clinical xenotrans-plantation: The next medical revolution? Lancet 2012, 379, 672–683. [Google Scholar] [CrossRef]
- Mansour, D.; Roberts, S.; Lee, M.; Shukrallah, B.; Whitson, B.A. The Role of Ex-vivo Lung Perfusion (EVLP) in Lung Trans-plantation. In Thoracic Surgery; Nistor, C.E., Tsui, S., Kirali, K., Nistor, C., Aresu, G., Kocher, G.J., Eds.; Springer: Cham, Switzerland, 2020; pp. 977–986. [Google Scholar]
- Leucht, W.; Gregor, G.; Stier, H. Das Miniaturschwein, Versuchs- und Modeltier in Medizin und Biologie; VEB Gustav Fischer: Jena, Germany, 1982; pp. 13–23. [Google Scholar]
- Schachler, K.; Minx, J.-O.; Sürie, C.; Distl, O.; Metzger, J. Genetic characterisation of Mini-LEWE as resource population for experimental research. Berl Münch Tierärtl Wochenschr 2020, 133. [Google Scholar] [CrossRef]
- Gregor, G. Das Miniaturschwein „Mini-LEWE“ und seine Eigenschaften. Monatsh Veterinarmed 1976, 31, 624–629. [Google Scholar]
- Morozov, V.A.; Plotzki, E.; Rotem, A.; Barkai, U.; Denner, J. Extended microbiological characterization of Göttingen minipigs: Porcine cytomegalovirus and other viruses. Xenotransplantation 2016, 23, 490–496. [Google Scholar] [CrossRef]
- Morozov, V.A.; Morozov, A.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Extended Microbiological Characterization of Göttingen Minipigs in the Context of Xenotransplantation: Detection and Vertical Transmission of Hepatitis E Virus. PLoS ONE 2015, 10, e0139893. [Google Scholar] [CrossRef] [Green Version]
- Denner, J.; Längin, M.; Reichart, B.; Krüger, L.; Fiebig, U.; Mokelke, M.; Radan, J.; Mayr, T.; Milusev, A.; Luther, F.; et al. Impact of porcine cytomegalovirus on long-term or-thotopic cardiac xenotransplant survival. Sci. Rep. 2020, 10, 17531. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Reduction of the survival time of pig xenotransplants by porcine cytomegalovirus. Virol. J. 2018, 15, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotzki, E.; Heinrichs, G.; Kubícková, B.; Ulrich, R.G.; Denner, J. Microbiological characterization of a newly established pig breed, Aachen Minipigs. Xenotransplantation 2016, 23, 159–167. [Google Scholar] [CrossRef]
- Morozov, V.A.; Heinrichs, G.; Denner, J. Effective Detection of Porcine Cytomegalovirus Using Non-Invasively Taken Samples from Piglets. Viruses 2017, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patience, C.; Takeuchi, Y.; Weiss, R.A. Infection of human cells by an endogenous retrovirus of pigs. Nat. Med. 1997, 3, 282–286. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Patience, C.; Magre, S.; Weiss, R.A.; Banerjee, P.T.; Le Tissier, P.; Stoye, J.P. Host range and interference studies of three classes of pig endogenous retrovirus. J. Virol. 1998, 72, 9986–9991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J.; Tönjes, R.R. Infection Barriers to Successful Xenotransplantation Focusing on Porcine Endogenous Retroviruses. Clin. Microbiol. Rev. 2012, 25, 318–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs. Viruses 2019, 12, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J.; Schuurman, K.J. High prevalence of recombinant porcine endogenous retroviruses (PERV-A/Cs) in minipigs: A review on origin and presence. Viruses 2021, 13, 1869. [Google Scholar] [CrossRef]
- Spizzo, T.; Denner, J.; Gazda, L.; Martin, M.; Nathu, D.; Scobie, L.; Takeuchi, Y. First update of the International Xenotransplan-tation Association consensus statement on conditions for undertaking clinical trials of porcine islet products in type 1 di-abetes--Chapter 2a: Source pigs-preventing xenozoonoses. Xenotransplantation 2016, 23, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Wei, H.-J.; Lin, L.; George, H.; Wang, T.; Lee, I.-H.; Zhao, H.-Y.; Wang, Y.; Kan, Y.; Shrock, E.; et al. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9. Science 2017, 357, 1303–1307. [Google Scholar] [CrossRef] [Green Version]
- Güell, M. Genome-Wide PERV Inactivation in Pigs Using CRISPR/Cas9. Springer Protoc. Handb. 2020, 2110, 139–149. [Google Scholar] [CrossRef]
- Scobie, L.; Denner, J.; Schuurman, H.-J. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9, editorial commentary. Xenotransplantation 2017, 24, e12363. [Google Scholar] [CrossRef]
- Gazda, L.S.; Collins, J.; Lovatt, A.; Holdcraft, R.W.; Morin, M.J.; Galbraith, D.; Graham, M.; Laramore, M.A.; MacLean, C.; Black, J.; et al. A comprehensive microbiological safety approach for agarose encapsulated porcine islets intended for clinical trials. Xenotransplantation 2016, 23, 444–463. [Google Scholar] [CrossRef]
- Hartline, C.B.; Conner, R.L.; James, S.H.; Potter, J.; Gray, E.; Estrada, J.; Tector, M.; Tector, A.J.; Prichard, M.N. Xenotransplantation panel for the detection of infectious agents in pigs. Xenotransplantation 2018, 25, e12427. [Google Scholar] [CrossRef] [Green Version]
- Noordergraaf, J.; Schucker, A.; Martin, M.; Schuurman, H.-J.; Ordway, B.; Cooley, K.; Sheffler, M.; Theis, K.; Armstrong, C.; Klein, L.; et al. Pathogen elimination and prevention within a regulated, Designated Pathogen Free, closed pig herd for long-term breeding and production of xenotransplantation materials. Xenotransplantation 2018, 25, e12428. [Google Scholar] [CrossRef] [Green Version]
- Busse, H. Über weitere Maßnahmen zur Züchtung eines kleinen Versuchsschweines für medizinische Zwecke. Monatsh Veterinarmed 1967, 23, 949–954. [Google Scholar]
- Schachler, K.; Suerie, C.; Minx, J.; Distl, O.; Metzger, J. Estimation of the genetic diversity in the Mini-Lewe breeding population and influence of inbreeding on litter traits. Zuchtungskd 2019, 91, 227–245. [Google Scholar]
- Yang, L.; Guell, M.; Niu, D.; George, H.; Lesha, E.; Grishin, D.; Aach, J.; Shrock, E.; Xu, W.; Poci, J. Genome-wide inacti-vation of porcine endogenous retroviruses (PERVs). Science 2015, 350, 1101–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieckhoff, B.; Kessler, B.; Jobst, D.; Kues, W.; Petersen, B.; Pfeifer, A.; Kurth, R.; Niemann, H.; Wolf, E.; Denner, J. Distri-bution and expression of porcine endogenous retroviruses in multi-transgenic pigs generated for xenotransplantation. Xenotransplantation 2009, 16, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.C.; Quinn, G.; Suling, K.M.; Oldmixon, B.A.; Van Tine, B.A.; Cina, R.; Arn, S.; Huang, C.A.; Scobie, L.; Onions, D.E.; et al. Identification of Exogenous Forms of Human-Tropic Porcine Endogenous Retrovirus in Miniature Swine. J. Virol. 2004, 78, 2494–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Groenke, N. Molecular Mechanism of Virus Attenuation by Codon Pair Deoptimization. Ph.D. Thesis, Free University, Berlin, Germany, 2019. [Google Scholar]
- Mueller, N.J.; Barth, R.; Yamamoto, S.; Kitamura, H.; Patience, C.; Yamada, K.; Cooper, D.K.C.; Sachs, D.H.; Kaur, A.; Fishman, J.A. Activation of Cytomegalovirus in Pig-to-Primate Organ Xenotransplantation. J. Virol. 2002, 76, 4866–4872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmielewicz, B.; Goltz, M.; Franz, T.; Bauer, C.; Brema, S.; Ellerbrok, H.; Beckmann, S.; Rziha, H.-J.; Lahrmann, K.-H.; Romero, C.; et al. A novel porcine gammaherpesvirus. Virology 2003, 308, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Xiao, Y.; Li, X.; Li, S.; Xie, N.; Yan, X.; Li, X.; Zhu, J. Development and application of a quadruplex real-time PCR assay for differential detection of porcine circoviruses (PCV1 to PCV4) in Jiangsu province of China from 2016 to 2020. Transbound. Emerg. Dis. 2021, 68, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2017, 91, e01879-16. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Shen, H.G.; Pal, N.; Ramamoorthy, S.; Huang, Y.W.; Lager, K.M.; Beach, N.M.; Halbur, P.G.; Meng, X. A Live-Attenuated Chimeric Porcine Circovirus Type 2 (PCV2) Vaccine Is Transmitted to Contact Pigs but Is Not Upregulated by Concurrent Infection with Porcine Parvovirus (PPV) and Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) and Is Efficacious in a PCV2b-PRRSV-PPV Challenge Model. Clin. Vaccine Immunol. 2011, 18, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Duvigneau, J.; Hartl, R.; Groiss, S.; Gemeiner, M. Quantitative simultaneous multiplex real-time PCR for the detection of porcine cytokines. J. Immunol. Methods 2005, 306, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Facci, M.R.; Auray, G.; Meurens, F.; Buchanan, R.; van Kessel, J.; Gerdts, V. Stability of expression of reference genes in porcine peripheral blood mononuclear and dendritic cells. Veter. Immunol. Immunopathol. 2011, 141, 11–15. [Google Scholar] [CrossRef] [PubMed]
- IDT. Tips for Working with gBlocks Gene Fragments—How to Resuspend, Quantify, and Calculate Copy Number. Available online: https://eu.idtdna.com/pages/education/decoded/article/tips-for-working-with-gblocks-gene-fragments (accessed on 30 September 2021).
- Zhang, H.H.; Hu, W.Q.; Li, J.Y.; Liu, T.N.; Zhou, J.Y.; Opriessnig, T.; Xiao, C.T. Novel circovirus species identified in farmed pigs designated as Porcine circovirus 4, Hunan province, China. Transbound. Emerg. Dis. 2019, 67, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.-T.; Giménez-Lirola, L.; Huang, Y.-W.; Meng, X.-J.; Halbur, P.G.; Opriessnig, T. The prevalence of Torque teno sus virus (TTSuV) is common and increases with the age of growing pigs in the United States. J. Virol. Methods 2012, 183, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, R.; Fiebig, U.; Norley, S.; Gürtler, L.; Kurth, R.; Denner, J. A neutralization assay for HIV-2 based on measurement of provirus integration by duplex real-time PCR. J. Virol. Methods 2009, 159, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Han, F.; Zou, Y.; Zhu, L.; Li, C.; Xu, Y.; Zhang, C.; Tan, F.; Wang, J.; Tao, S.; et al. Rapid detection of porcine circovirus type 2 using a TaqMan-based real-time PCR. Virol. J. 2010, 7, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, K.J.; Minihan, D.; Campion, E.M.; Loughran, S.T.; Allan, G.; McNeilly, F.; Walls, D. Infection of pigs in Ireland with lymphotropic γ-herpesviruses and relationship to postweaning multisystemic wasting syndrome. Veter. Microbiol. 2006, 116, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.; Benes, V.; Garson, J.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.; Shipley, G.; et al. The need for transparency and good practices in the qPCR literature. Nat. Methods 2013, 10, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Becker, A.-C.; Kraemer, A.; Epe, C.; Strube, C. Sensitivity and efficiency of selected coproscopical methods—sedimentation, combined zinc sulfate sedimentation-flotation, and McMaster method. Parasitol. Res. 2016, 115, 2581–2587. [Google Scholar] [CrossRef]
- Gregor, G.; H-Bohrisch, H.-G.; Knorck, H. Erfahrungen mit der spezifisch-pathogenfreien Haltung (SPF) des Miniaturschweines Mini-Lewe (Experience with specific-pathogen-free (SPF) "Mini-Lewe" miniature pigs). MH Vet. Med. 1980, 35, 190–193. [Google Scholar]
- Denner, J. Sensitive detection systems for infectious agents in xenotransplantation. Xenotransplantation 2020, e12594. [Google Scholar] [CrossRef] [Green Version]
- Denner, J. Xenotransplantation and Hepatitis E virus. Xenotransplantation 2015, 22, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Pischke, S.; Steinmann, E.; Blümel, J.; Glebe, D. Why all blood donations should be tested for hepatitis E virus (HEV). BMC Infect. Dis. 2019, 19, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Krüger, L.; Längin, M.; Reichart, B.; Fiebig, U.; Kristiansen, Y.; Prinz, C.; Kessler, B.; Egerer, S.; Wolf, E.; Abicht, J.-M.; et al. Transmission of Porcine Circovirus 3 (PCV3) by Xenotransplantation of Pig Hearts into Baboons. Viruses 2019, 11, 650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynyard, S.; Nathu, D.; Garkavenko, O.; Denner, J.; Elliott, R. Microbiological safety of the first clinical pig islet xenotransplantation trial in New Zealand. Xenotransplantation 2014, 21, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Wynyard, S.; Matsumoto, S.; Abalovich, A.; Denner, J.; Elliott, R. No PERV transmission during a clinical trial of pig islet cell transplantation. Virus Res. 2017, 227, 34–40. [Google Scholar] [CrossRef]
- Paradis, K.; Langford, G.; Long, Z.; Heneine, W.; Sandstrom, P.; Switzer, W.M.; Chapman, L.E.; Lockey, C.; Onions, D.; Otto, E. Search for Cross-Species Transmission of Porcine Endogenous Retrovirus in Patients Treated with Living Pig Tissue. Science 1999, 285, 1236–1241. [Google Scholar] [CrossRef] [Green Version]
- Denner, J.; Schuurman, H.J.; Patience, C. The International Xenotransplantation Association consensus statement on conditions for undertaking clinical trials of porcine islet products in type 1 diabetes--chapter 5: Strategies to prevent transmission of porcine endogenous retroviruses. Xenotransplantation 2009, 16, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Abicht, J.M.; Reichart, B.; Mayr, T.; Guethoff, S.; Denner, J. Active replication of porcine cytomegalovirus (PCMV) following transplantation of a pig heart into a baboon despite undetected virus in the donor pig. Ann. Virol. Res. 2016, 2, 1018. [Google Scholar]
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Créhan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; Zhu, J.; et al. Coinfections and their molecular consequences in the porcine respiratory tract. Veter. Res. 2020, 51, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.I.; Wilkinson, R.; Fishman, J.A. Genomic presence of recombinant porcine endogenous retrovirus in transmitting miniature swine. Virol. J. 2006, 3, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Fishman, J.A.; Sachs, D.H.; Yamada, K.; Wilkinson, R.A. Absence of interaction between porcine endogenous retrovirus and porcine cytomegalovirus in pig-to-baboon renal xenotransplantation in vivo. Xenotransplantation 2018, 25, e12395. [Google Scholar] [CrossRef]
- Yamada, K.; Tasaki, M.; Sekijima, M.; Wilkinson, R.A.; Villani, V.; Moran, S.G.; Cormack, T.A.; Hanekamp, I.; Arn, J.S.; Fishman, J.A.; et al. Porcine Cytomegalovirus Infection Is Associated With Early Rejection of Kidney Grafts in a Pig to Baboon Xenotransplantation Model. Transplantation 2014, 98, 411–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittmann, I.; Mihica, D.; Plesker, R.; Denner, J. Expression of porcine endogenous retroviruses (PERV) in different organs of a pig. Virology 2012, 433, 329–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieckhoff, B.; Puhlmann, J.; Büscher, K.; Hafner-Marx, A.; Herbach, N.; Bannert, N.; Büttner, M.; Wanke, R.; Kurth, R.; Denner, J. Expression of porcine endogenous retroviruses (PERVs) in melanomas of Munich miniature swine (MMS) Troll. Veter. Microbiol. 2007, 123, 53–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [Green Version]
- Kijlstra, A.; Meerburg, B.; Mul, M. Animal-friendly production systems may cause re-emergence of Toxoplasma gondii. NJAS Wagening. J. Life Sci. 2004, 52, 119–132. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Sample ID | Gender | Animal Status | Age at the Time of Sampling |
|---|---|---|---|
| LEWE-1 | female | breeding animal | 10 years |
| LEWE-2 | female | breeding animal | 6 years |
| LEWE-3 | female | breeding animal | 5 years |
| LEWE-4 | male | breeding animal | 6 months |
| LEWE-5 | male | offspring | 13 months |
| LEWE-6 | male | offspring | 13 months |
| LEWE-7 | male | offspring | 6 months |
| LEWE-8 | male | offspring | 6 months |
| LEWE-9 | male | offspring | 4 months |
| LEWE-10 | male | offspring | 4 months |
| PCR Assay | Primer/Probe | Sequence 5′-3′ | Amplicon (Base Pair) | Reference |
|---|---|---|---|---|
| HEV | JVHEV-Fwd | GGT GGT TTC TGG GGT GAC | 70 | [39] |
| JVHEV-Rev | AGG GGT TGG TTG GAT GAA | |||
| JVHEV-Probe | 6FAM-TGA TTC TCA GCC CTT CGC-BHQ | |||
| PCMV | PCMV-Fwd | ACT TCG TCG CAG CTC ATC TGA | 63 | [41] modified |
| PCMV-Rev | GTT CTG GGA TTC CGA GGT TG | |||
| PCMV-Probe | 6FAM-CAG GGC GGC GGT CGA GCT C-BHQ | |||
| PLHV-1 | PLHV-1 (1125)-Fwd | CTC ACC TCC AAA TAC AGC GA | 73 | [42] |
| PLHV-1 (1125)-Rev | GCT TGA ATC GTG TGT TCC ATA G | |||
| PLHV-1 (1125)-Probe | 6FAM-CTG GTC TAC TGA ATC GCC GCT AAC AG-TAMRA | |||
| PLHV-2 | PLHV-2 (1155)-Fwd | GTC ACC TGC AAA TAC ACA GG | 74 | [42] |
| PLHV-2 (1155)-Rev | GGC TTG AAT CGT ATG TTC CAT AT | |||
| PLHV-2 (1155)-Probe | 6FAM-CTG GTC TAC TGA AGC GCT GCC AAT AG-TAMRA | |||
| PLVH-3 | PLHV-3 (1156)-Fwd | CTC ACC TCC AAA TAC AGC GA | 73 | [42] |
| PLHV-3 (1156)-Rev | GCT TGA ATC GTG TGT TCC ATA G | |||
| PLHV-3 (1156)-Probe | 6FAM-CTGGTCTACTGAATCGCCGCTAACAG-TAMRA | |||
| PCV1 | PCV-1 (F2020)-Fwd | AAC CCC ATA AGA GGT GGG TGT T | 129 | [43] modified |
| PCV-1 (F2020)-Rev | TTC TAC CCT CTT CCA AAC CTT CCT | |||
| PCV-1 (F2020)-Probe | 6FAM-TCC GAG GAG GAG AAA AAC AAA ATA CGGGA-BHQ1 | |||
| PCV2 | PCV-2 (F2020)-Fwd | CTG AGT CTT TTT TAT CAC TTC GTA ATG GT | 146 | [43] modified |
| PCV-2 (F2020)-Rev | ACT GCG TTC GAA AAC AGT ATA TAC GA | |||
| PCV-2 (F2020)-Probe | 6FAM-TTA AGT GGG GGG TCT TTA AGA TTA AAT TCT CTG AAT TGT-TAMRA | |||
| PCV3 | PCV-3-Fwd | AGT GCT CCC CAT TGA ACG | 112 | [44] |
| PCV-3-Rev | ACA CAG CCG TTA CTT CAC | |||
| PCV-3-Probe | 6FAM-ACC CCA TGG CTC AAC ACA TAT GAC C-BHQ1 | |||
| PCV4 | PCV-4 (F2020)-Fwd | ATT ATT AAA CAG ACT TTA TTT GTG TCA TCA CTT | 103 | [43] |
| PCV-4 (F2020)-Rev | ACA GGG ATA ATG CGT AGT GAT CAC T | |||
| PCV-4 (F2020)-Probe | 6FAM-ATA CTA CAC TTG ATC TTA GCC AAA AGG CTC GTT GA-BHQ1 | |||
| PPV-1 | PPV-1-Fwd | CAG AAT CAG CAA CCT CAC CA | 106 | [45] modified |
| PPV-1-Rev | GCT GCT GGT GTG TAT GGA AG | |||
| PPV-1-Probe | 6FAM-TGC AAG CTT/ZEN/AAT GGT CGC ACT AGA CA-BHQ1 | |||
| PERVpol | PERVpol-Fwd | CGA CTG CCC CAA GGG TTC AA | 236 | [36] |
| PERVpol-Rev | TCT CTC CTG CAA ATC TGG GCC | |||
| PERVpol-Probe | 6FAM-CAC GTA CTG GAG GAG GGT CAC CTG-BHQ1 | |||
| PERV-C | envC.2-Fwd | GAT TAG AAC TGG AAG CCC CAA GTG CTC T | 288 | [37] |
| envC.2-Rev | TCT GAT CCA GAA GTT ATG TTA GAG GAT GGT | |||
| PERV-A/C | PERV-A env VRBF-Fwd | CCT ACC AGT TAT AAT CAA TTT AAT TAT GGC | 1266 | [38] |
| PERV-C env TMR-Rev | CTC AAA CCA CCC TTG AGT AGT TTC C | |||
| pGAPDH | pGAPDH-Fwd | ACA TGG CCT CCA AGG AGT AAG A | 106 | [46] |
| pGAPDH-Rev | GAT CGA GTT GGG GCT GTG ACT | |||
| pGAPDH-Probe | HEX-CCA CCA ACC CCA GCA AGA GCA CGC-BHQ1 | |||
| Flu-A (NA) | NG05 NA-Fwd | CTG GAC TAG TGG GAG CAT CA | 93 | [40] modified |
| NG06 NA-Rev | ATG GTG AAC GGC AAC TCA G | |||
| NG07 NA-Probe | HEX-CAC CGT CTG GCC AAG ACC AAT C-BHQ1 |
| Sample ID | PERVpol | PERV-C | PERV-A/C |
|---|---|---|---|
| LEWE-1 | 15.12 | positive | negative |
| LEWE-2 | 14.47 | positive | negative |
| LEWE-3 | 15.37 | positive | negative |
| LEWE-4 | 16.14 | positive | negative |
| LEWE-5 | 14.01 | positive | negative |
| LEWE-6 | 15.31 | positive | negative |
| LEWE-7 | 15.43 | positive | negative |
| LEWE-8 | 17.49 | positive | negative |
| LEWE-9 | 16.01 | positive | negative |
| LEWE-10 | 16.70 | positive | negative |
| Sample ID | HEV | Flu-A (NA) |
|---|---|---|
| LEWE-1 | no Ct | 27.20 * |
| LEWE-2 | no Ct | 26.59 |
| LEWE-3 | no Ct | 26.83 |
| LEWE-4 | no Ct | 27.23 |
| LEWE-5 | no Ct | 32.91 |
| LEWE-6 | no Ct | 31.06 |
| LEWE-7 | no Ct | 31.83 |
| LEWE-8 | no Ct | 27.60 |
| LEWE-9 | no Ct | 32.17 |
| LEWE-10 | no Ct | 26.92 |
| Minipig Breed 1 | Institution/ Company | PERV-C | PERV-A/C | PCMV | PCV1 | PCV2 | PCV3 | HEV | PLHV-1 | PLHV-2 | PLHV-3 | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mini LEWE 2 | University of Veterinary Medicine Hannover, Germany | 10/10 PBMCs | 0/10 PBMCs | 0/10 PBMCs | 0/10 PBMCs | 0/10 PBMCs | 0/10 PBMCs | 0/10 PBMCs | 0/10 PBMC | 0/10 PBMCs | 0/10 PBMCs | this manuscript |
| Göttingen minipigs | Ellegaard Göttingen Minipigs A/S, Denmark | 28/28 | None PBMCs | 10/22 PBMC liver spleen | n.t. 3 | 3/21 PBMC liver spleen | 0/10 PBMC liver spleen | 9/40 serum | 1/10 anti-bodies | 0/5 PBMC liver spleen | 0/5 PBMC liver spleen | [10,16,17] |
| Göttingen minipigs | University Göttingen, Göttingen, Germany | 11/11 | 3/13 PBMCs | PBMCs 0/10 | n.t. | 3/10 PBMCs | 0/10 PBMCs | 0/10 PBMCs | 2/11 PBMCs | 2/11 PBMCs | 2/11 PBMCs | [25] |
| Aachen minipigs 4 | Aachen Minipig, Heinsberg, Germany | 18/18 | 3/18 liver spleen | 5/18 PBMCs | n.t. | 6/18 antibodies | n.t. | 12/18 serum | 0/18 PBMCs | 5/18 PBMCs | 2/18 PBMCs | [20] |
| Yucatan micropig | Charles River, Saint-Aubin-Les-Elbeuf, France | 1/1 5 | 1/1 PBMCs | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [72] |
| Munich miniature swine (MMS) 6 | Institute of Veterinary Pathology, University of Munich | Yes | 5/5 PBMCs | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halecker, S.; Metzger, J.; Strube, C.; Krabben, L.; Kaufer, B.; Denner, J. Virological and Parasitological Characterization of Mini-LEWE Minipigs Using Improved Screening Methods and an Overview of Data on Various Minipig Breeds. Microorganisms 2021, 9, 2617. https://doi.org/10.3390/microorganisms9122617
Halecker S, Metzger J, Strube C, Krabben L, Kaufer B, Denner J. Virological and Parasitological Characterization of Mini-LEWE Minipigs Using Improved Screening Methods and an Overview of Data on Various Minipig Breeds. Microorganisms. 2021; 9(12):2617. https://doi.org/10.3390/microorganisms9122617
Chicago/Turabian StyleHalecker, Sabrina, Julia Metzger, Christina Strube, Ludwig Krabben, Benedikt Kaufer, and Joachim Denner. 2021. "Virological and Parasitological Characterization of Mini-LEWE Minipigs Using Improved Screening Methods and an Overview of Data on Various Minipig Breeds" Microorganisms 9, no. 12: 2617. https://doi.org/10.3390/microorganisms9122617